La teoría de Mendel explica por qué algunos rasgos pasan de padres a hijos de forma bastante predecible y por qué otros no obedecen a reglas tan simples. Es la base de la herencia genética clásica: habla de genes en pares, alelos dominantes y recesivos, y de cómo se reparten durante la formación de gametos. En genética médica sigue siendo una referencia útil, pero solo funciona bien cuando el rasgo encaja de verdad en ese modelo.
Lo esencial para entender la herencia mendeliana
- Un rasgo mendeliano suele depender de un solo gen con dos alelos principales.
- Las tres ideas clave son dominancia, segregación e independencia de caracteres.
- Los cruces clásicos producen proporciones como 3:1 o 9:3:3:1, pero no siempre en humanos.
- Muchos rasgos comunes son poligénicos y ya no siguen un patrón mendeliano puro.
- En medicina personalizada, este modelo ayuda a interpretar árboles familiares y resultados genéticos.
Qué explica la herencia mendeliana
Yo suelo empezar por una idea simple: heredamos dos copias de casi cada gen, una de cada progenitor. A esas variantes se las llama alelos, y su combinación forma el genotipo; lo que vemos en el cuerpo, como color de semilla o una característica clínica, es el fenotipo. Mendel trabajó con guisantes porque le permitían observar rasgos muy claros y contar resultados con precisión, algo que hoy sigue siendo una lección metodológica muy útil.
En ese modelo, un alelo puede ser dominante o recesivo. Dominante no significa “mejor” ni “más frecuente”; significa que basta una copia para que el rasgo se exprese. Recesivo, en cambio, suele necesitar dos copias para manifestarse. Esa diferencia es sencilla sobre el papel, pero cambia por completo la forma de leer una familia, un árbol genealógico o un informe genético.
Para no mezclar conceptos, yo recomiendo quedarse con este mapa mental: alelo es la versión del gen, genotipo es la combinación heredada y fenotipo es el resultado observable. Cuando esos tres niveles se entienden bien, el resto de la teoría se vuelve mucho más lógico. Y precisamente por eso conviene separar ahora las leyes que Mendel formuló a partir de sus cruces.
Las tres leyes que ordenan el modelo clásico
Las leyes de Mendel no son dogmas cerrados; son una forma muy eficaz de describir cómo se comportan ciertos genes cuando la herencia es relativamente simple. En la práctica, el valor del modelo está en que permite prever proporciones y detectar cuándo un patrón familiar tiene sentido y cuándo no.
| Ley | Qué dice | Ejemplo breve | Cuándo se complica |
|---|---|---|---|
| Uniformidad o dominancia | Al cruzar dos líneas puras distintas, la primera generación suele ser uniforme y muestra el rasgo dominante. | TT × tt produce descendencia Tt con fenotipo alto. | Cuando hay dominancia incompleta o codominancia. |
| Segregación | Las dos copias de un gen se separan al formar gametos, de modo que cada gameto recibe solo una. | Un progenitor Tt puede formar gametos T o t. | Cuando intervienen más genes, penetrancia incompleta o mosaicos especiales. |
| Distribución independiente | Los genes de rasgos distintos se heredan de manera independiente si no están ligados. | La forma de una semilla no determina su color. | Cuando los genes están cerca en el mismo cromosoma. |
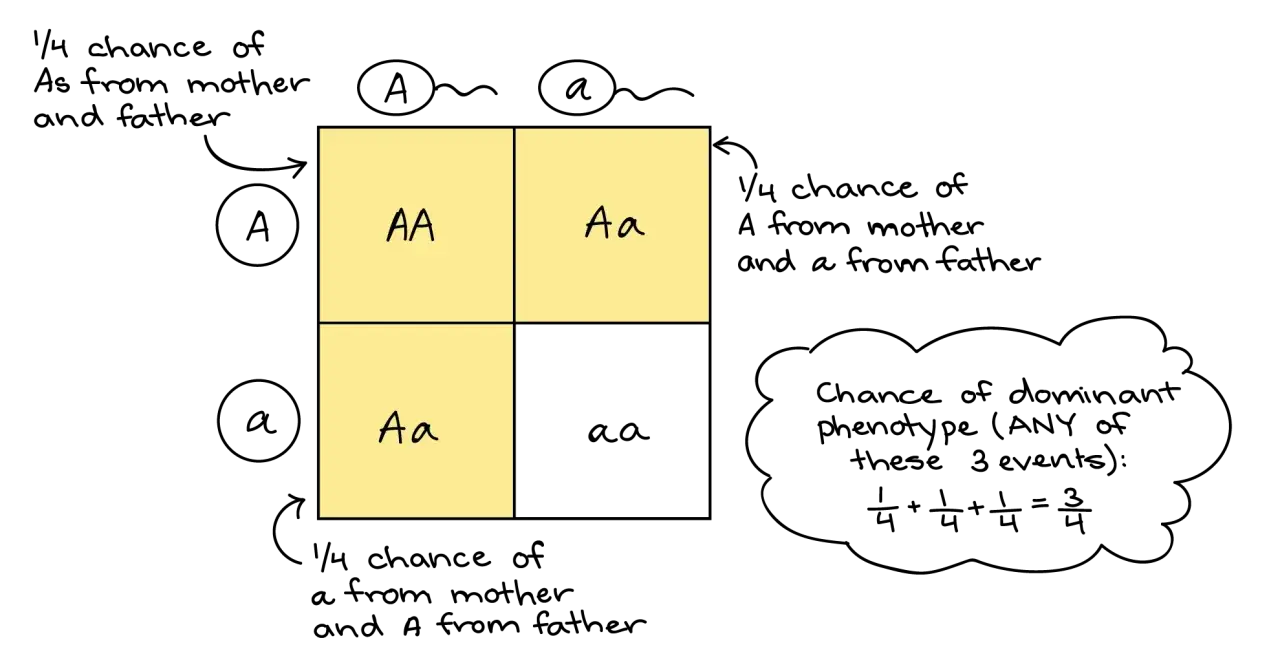
La consecuencia más famosa de este esquema es la proporción 3:1 en un cruce monohíbrido y 9:3:3:1 en uno dihíbrido. Pero yo siempre matizo lo mismo: esas proporciones son una guía, no una promesa universal. Funcionan muy bien en los ejemplos de libro, y muchas veces también en genética clínica, aunque no describen toda la biología real.
Con esto ya se entiende lo más importante del modelo. El siguiente paso es verlo funcionando en un caso concreto, porque ahí es donde las ideas dejan de sonar abstractas y empiezan a servir de verdad.
Un cruce sencillo ayuda más que una definición larga
Imaginemos una planta con semillas amarillas y otra con semillas verdes. Si la variedad amarilla es dominante y ambas líneas son puras, la primera generación tendrá semillas amarillas aunque una de las copias sea “verde” en silencio. Eso ya muestra una de las intuiciones más potentes de Mendel: los rasgos no se mezclan como pintura, sino que se transmiten como unidades discretas.
Si cruzamos después dos individuos de la primera generación, obtenemos una segunda generación con genotipos en proporción 1:2:1 y fenotipos en proporción 3:1. Traducido a lenguaje llano: una parte será homocigota dominante, dos partes heterocigotas y una parte homocigota recesiva. Esa lógica sigue siendo muy útil para entender por qué un rasgo recesivo puede “saltarse” una generación.
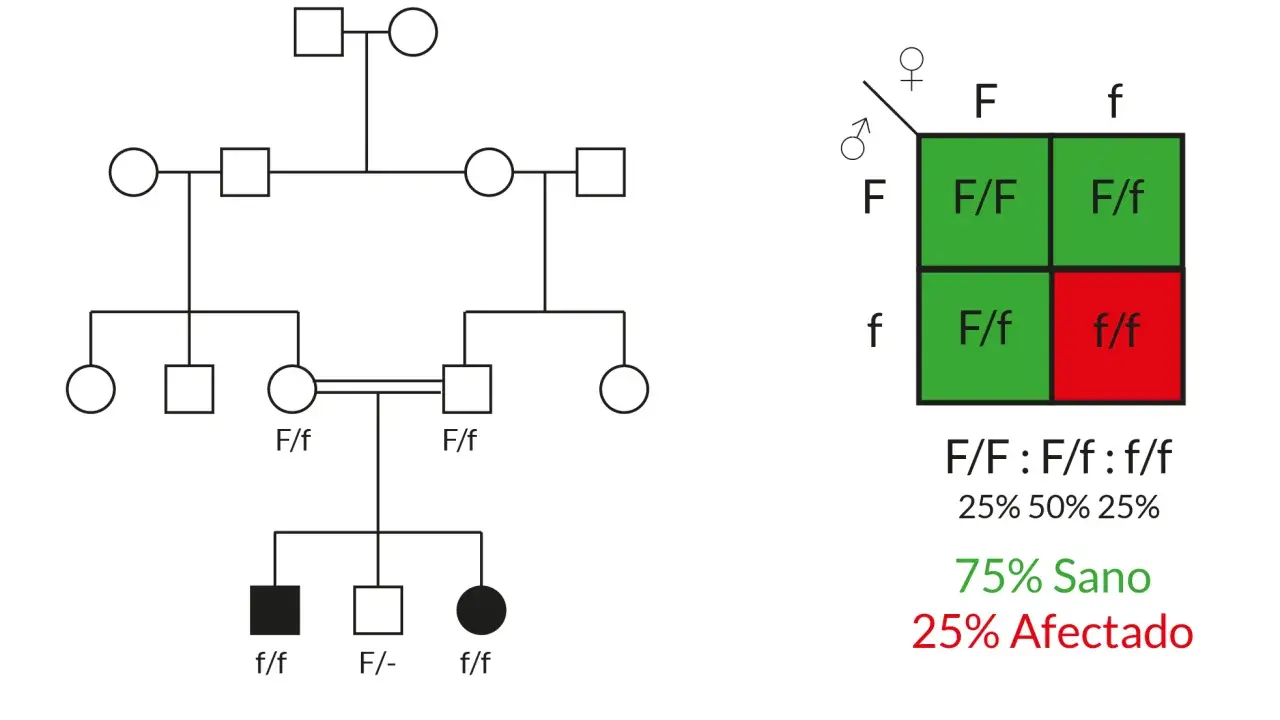
En humanos, el esquema funciona bien con algunas enfermedades monogénicas, por ejemplo cuando una variante concreta basta para explicar gran parte del cuadro o cuando dos copias alteradas son necesarias para que aparezca la enfermedad. Aun así, yo no lo usaría nunca como atajo diagnóstico: en personas reales hay penetrancia variable, historia clínica, edad de inicio y contexto familiar. El patrón heredado da pistas, pero no reemplaza la interpretación clínica.
Y justo ahí empiezan los matices importantes, porque muchos rasgos no encajan en el modelo clásico aunque durante años se hayan explicado mal de forma demasiado simple.
Dónde el modelo deja de ser suficiente
El NHGRI recuerda que los rasgos poligénicos, como la estatura o el color de la piel, no siguen patrones de herencia mendeliana. Esa precisión importa mucho, porque en biología es fácil confundir “heredable” con “mendeliano”, y no son lo mismo. Hay rasgos que sí se heredan, pero lo hacen bajo una arquitectura más compleja.
| Situación | Qué cambia | Cómo se ve en la práctica |
|---|---|---|
| Dominancia incompleta | El heterocigoto muestra un fenotipo intermedio. | El rasgo no se ve “apagado” ni completamente dominante. |
| Codominancia | Las dos variantes se expresan a la vez. | El sistema ABO es el ejemplo clásico que muchos estudiantes reconocen enseguida. |
| Ligamiento génico | Genes cercanos en el mismo cromosoma tienden a heredarse juntos. | La independencia entre rasgos deja de cumplirse de forma limpia. |
| Rasgos poligénicos | Intervienen muchos genes a la vez. | La estatura, parte de la pigmentación o la presión arterial no se explican con un solo gen. |
| Herencia ligada al sexo o mitocondrial | La transmisión depende del cromosoma X/Y o de la línea materna. | El patrón familiar cambia mucho respecto al esquema autosómico clásico. |
Además de estas situaciones, hay otro matiz que me parece especialmente importante: penetrancia y expresividad. La penetrancia indica si una variante se manifiesta o no, y la expresividad, cuánto y cómo se manifiesta. Dos personas con la misma variante pueden mostrar cuadros muy distintos, y eso rompe la idea ingenua de que un gen equivale siempre a un efecto fijo.
La lectura útil no es pensar que Mendel “se equivocó”, sino entender que describió una parte del problema. Hoy sabemos que la herencia es más rica, más desigual y, en algunos rasgos, bastante más difícil de resumir en un cruce escolar. Esa es precisamente la razón por la que su trabajo sigue enseñándose: porque sirve de base para notar cuándo la realidad se aparta del patrón sencillo.
Por qué sigue siendo útil en genética médica y medicina personalizada
En consulta genética, yo veo el modelo mendeliano como una primera capa de orden. Si un rasgo sigue una herencia dominante, recesiva o ligada al X, el árbol familiar puede orientar de inmediato el riesgo de recurrencia, el estudio de portadores y la lectura de un resultado molecular. Sin esa base, un informe genético se interpreta peor y se mezclan hipótesis que no encajan entre sí.
También es útil para distinguir entre una variante con gran efecto y un rasgo complejo. No es lo mismo encontrar una mutación patogénica en un gen que explica casi todo el cuadro, que encontrar muchas variantes pequeñas que solo aumentan algo la predisposición. En medicina personalizada esa diferencia cambia decisiones: desde el consejo genético hasta la priorización de pruebas o el valor real de un panel multigénico.
Otro uso práctico está en la interpretación de familiares no afectados. Un caso recurrente es el de los portadores sanos en enfermedades recesivas: pueden no tener síntomas, pero sí transmitir la variante. Eso obliga a pensar en probabilidades, no en certezas absolutas. Y obliga también a leer bien el término “portador”, que en genética clínica tiene un significado preciso y no debería usarse como sinónimo de enfermedad.
En otras palabras, Mendel no compite con la secuenciación masiva ni con los paneles de genes; los ordena. La tecnología detecta variantes, pero el patrón de herencia sigue siendo la forma más rápida de preguntar si esas variantes explican de verdad lo que vemos en la familia. Sin ese filtro, es fácil sobrerinterpretar hallazgos que en realidad tienen un peso menor.
Con esa idea en mente, el cierre lógico es repasar qué conviene revisar antes de dar por hecho que un rasgo sigue un patrón mendeliano puro.
Qué conviene revisar antes de dar un rasgo por mendeliano
- Si el rasgo es cualitativo o cuantitativo: los rasgos continuos suelen ser poligénicos.
- Si aparece en casi todas las generaciones o si salta algunas: eso orienta la herencia dominante o recesiva.
- Si afecta por igual a hombres y mujeres: un sesgo por sexo puede apuntar a herencia ligada al X o a otra explicación.
- Si el fenotipo se expresa siempre de la misma manera: la variabilidad puede deberse a penetrancia incompleta o expresividad variable.
- Si hay varios genes implicados o una influencia ambiental clara: en ese caso el patrón simple pierde fuerza.
- Si alguien confunde dominante con frecuente o recesivo con “leve”: ese error lleva a interpretar mal familias enteras.
Yo me quedaría con una idea práctica muy concreta: el valor de la herencia mendeliana no está en memorizar proporciones, sino en saber leer patrones. Cuando un rasgo encaja, Mendel sigue siendo una herramienta limpia y poderosa; cuando no encaja, la biología moderna ofrece mejores explicaciones que forzar el esquema clásico. Esa es la forma más honesta y útil de entender la genética hereditaria hoy.