La copia del material genético no funciona como una fotocopia literal, sino como un sistema coordinado que abre la doble hélice, lee cada hebra y corrige errores en tiempo real. Entender la replicación del ADN ayuda a explicar cómo se dividen las células, por qué aparecen mutaciones y qué ocurre cuando fallan los mecanismos de reparación. Yo la considero una de las piezas más importantes de la biología molecular porque conecta genética básica, cáncer y medicina personalizada.
Lo esencial de este proceso en pocas líneas
- Es un proceso semiconservativo: cada molécula hija conserva una hebra original y sintetiza una nueva.
- Comienza en orígenes de replicación y avanza formando horquillas de replicación.
- La ADN polimerasa solo construye en dirección 5'→3', así que una hebra se copia de forma continua y la otra en fragmentos.
- La corrección de errores es parte del proceso: el proofreading y la reparación posterior reducen mucho los fallos.
- En eucariotas hay una dificultad extra: los cromosomas son lineales y los telómeros exigen un control adicional.
Cómo se copia el ADN paso a paso
Si tuviera que resumir el mecanismo en una sola idea, diría que la célula hace tres cosas en orden: abre la doble hélice, copia la información y verifica el resultado. Ese movimiento ocurre en la horquilla de replicación, una estructura en forma de Y donde se organiza toda la maquinaria enzimática.
Inicio
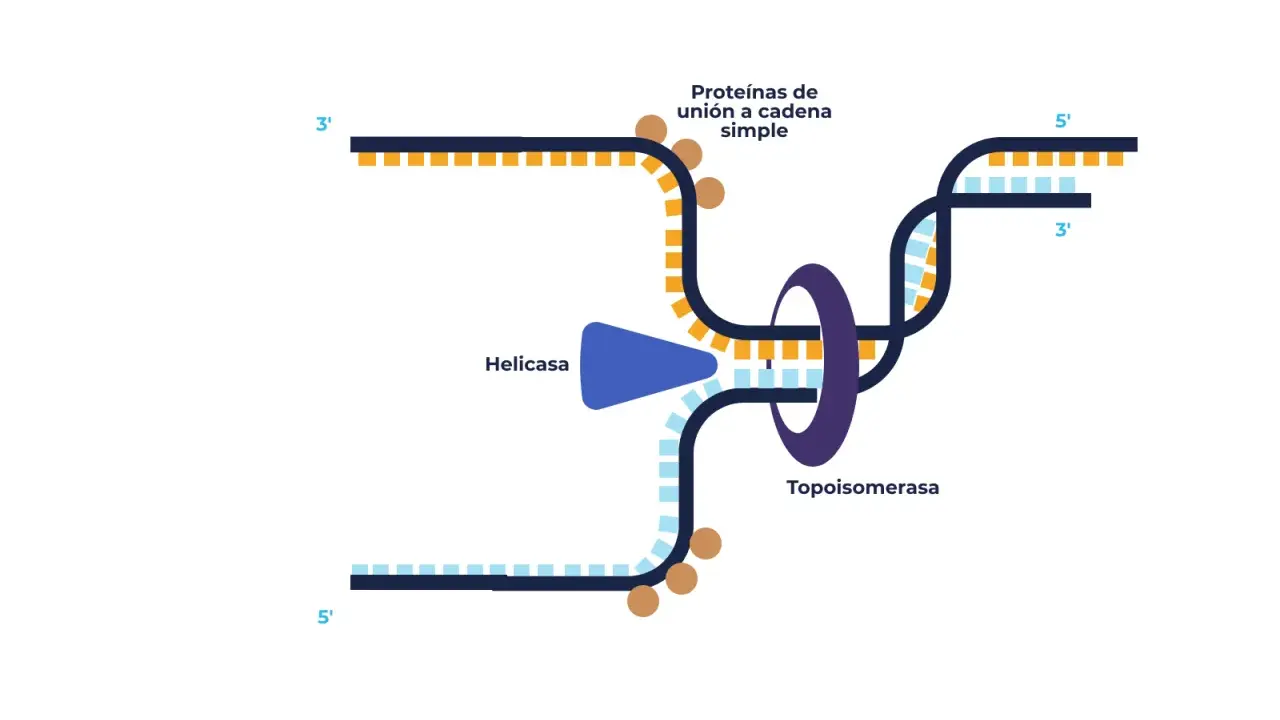
La replicación arranca en regiones concretas del ADN llamadas orígenes de replicación. Allí se ensamblan proteínas iniciadoras que reclutan a la helicasa, la enzima que separa las dos hebras rompiendo los puentes de hidrógeno entre las bases. En eucariotas no suele haber un único punto de arranque, sino miles de orígenes distribuidos por el genoma, porque copiar cromosomas grandes exige varios frentes de trabajo al mismo tiempo.
Elongación
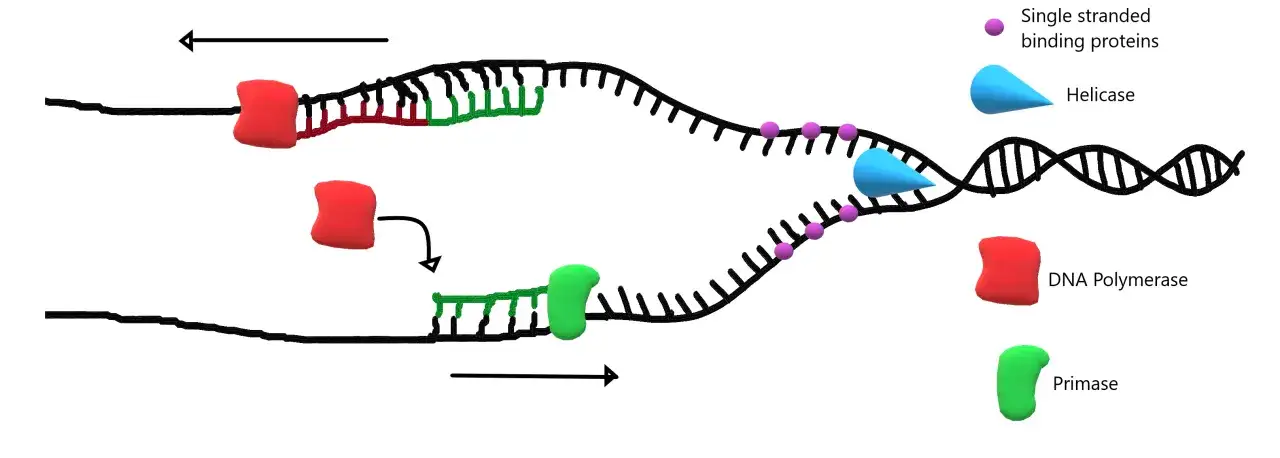
Una vez abierta la doble hélice, la ADN polimerasa comienza a añadir nucleótidos complementarios sobre la hebra molde. Aquí aparece una regla que conviene no olvidar: la síntesis siempre va en dirección 5'→3'. Eso obliga a distinguir entre la hebra líder, que se copia de forma continua, y la hebra rezagada, que se copia en fragmentos de Okazaki, pequeños segmentos que después se unirán entre sí.
La polimerasa no puede empezar sola desde cero. Necesita un cebador, normalmente un corto fragmento de ARN fabricado por la primasa, que le aporte el extremo 3'-OH libre para arrancar. Este detalle técnico explica buena parte de la arquitectura del proceso y por qué la replicación no es un simple “copiar y pegar”.
Terminación
La copia termina cuando las horquillas se encuentran o cuando el tramo correspondiente queda completamente duplicado. En ese punto, otras proteínas sellan las discontinuidades y restauran la continuidad del ADN. La ligasa, por ejemplo, une los fragmentos de Okazaki dejando una molécula completa y funcional.
Con esa secuencia clara, ya se entiende mejor por qué hace falta tanta proteína auxiliar. La siguiente pieza del rompecabezas es precisamente esa maquinaria de apoyo.
Las enzimas que sostienen la copia fiel
Yo suelo separar las proteínas de replicación en cuatro grupos: las que abren, las que inician, las que alargan y las que estabilizan o rematan. Esa división ayuda mucho a no perderse entre nombres que, a primera vista, parecen una lista interminable.
| Componente | Función | Por qué importa |
|---|---|---|
| Helicasa | Separa las dos hebras de ADN | Abre la doble hélice y crea la horquilla de replicación |
| Primasa | Sintetiza un cebador corto de ARN | Permite que la ADN polimerasa empiece a copiar |
| ADN polimerasa | Añade nucleótidos complementarios y corrige parte de los errores | Es la enzima central de la copia del genoma |
| Topoisomerasa | Reduce la tensión y el superenrollamiento delante de la horquilla | Evita que el ADN se rompa o se bloquee por torsión excesiva |
| Proteínas SSB o RPA | Estabilizan el ADN monocatenario | Impedir que las hebras separadas vuelvan a unirse de forma desordenada |
| Ligasa | Sella los enlaces entre fragmentos vecinos | Completa la hebra rezagada y deja la cadena continua |
| PCNA o abrazadera deslizante | Retiene a la polimerasa unida al ADN | Aumenta la procesividad, es decir, la capacidad de copiar muchos nucleótidos sin soltarse |
La idea práctica aquí es sencilla: ninguna enzima trabaja sola. El replisoma es un complejo coordinado, y si una pieza falla, el resto se resiente. Por eso la replicación no se entiende bien si solo memorizas nombres; hay que ver qué papel cumple cada uno dentro del conjunto.
Y precisamente porque el sistema es tan fino, también necesita varias capas de control para evitar que un error puntual se convierta en una mutación estable.
Dónde fallan las copias y cómo lo corrige la célula
Incluso en un sistema muy afinado, el error cero no existe. Lo importante es que la célula combina varias barreras para que la mayoría de los fallos no lleguen a fijarse en el genoma.
Proofreading o corrección inmediata
Muchas ADN polimerasas tienen actividad exonucleasa 3'→5', que les permite retroceder y retirar el nucleótido incorrecto antes de seguir avanzando. Dicho de forma simple: la enzima no solo escribe, también relee una parte del texto mientras lo escribe.
Lee también: ARN - ¿Qué es y por qué es clave en genética y medicina?
Reparación de desajustes
Los errores que escapan al proofreading pueden corregirse después mediante la reparación de desajustes o mismatch repair, un sistema que reconoce bases mal apareadas y repara la hebra recién sintetizada. Esta capa es clave porque, en un genoma humano de unos 3.200 millones de pares de bases, un fallo aislado puede parecer pequeño, pero no lo es si se repite miles de veces.
Cuando un error no se corrige antes de la siguiente ronda de copia, se fija como mutación. A partir de ahí, puede no pasar nada, alterar la función de un gen o contribuir a una enfermedad si afecta a una región relevante. Por eso la fidelidad de la replicación es tan importante: no solo evita fallos, también preserva la estabilidad genética a largo plazo.
Ese margen de error aceptable cambia bastante según el tipo de célula, y ahí es donde las diferencias entre bacterias y células humanas se vuelven realmente interesantes.
Qué cambia entre bacterias y células humanas
La química básica es la misma, pero el contexto celular no lo es. En bacterias, el proceso suele ser más compacto; en eucariotas, la cromatina y los telómeros añaden capas de regulación que hacen la copia más compleja de montar y coordinar.
| Aspecto | Bacterias | Eucariotas | Por qué importa |
|---|---|---|---|
| Forma del cromosoma | Habitualmente circular | Lineal y repartido en varios cromosomas | Los extremos lineales plantean el problema de los telómeros |
| Orígenes de replicación | Normalmente uno principal por cromosoma | Miles de orígenes | Permite copiar genomas mucho más grandes en un tiempo razonable |
| Empaquetamiento | Más simple | ADN enrollado en nucleosomas y cromatina | Antes de copiar hay que abrir y después volver a organizar la estructura |
| Telomerasa | No suele ser necesaria | Activa en células germinales, algunas células madre y muchos tumores | Resuelve el problema de los extremos cromosómicos |
| Complejidad regulatoria | Más directa | Más capas de control ligadas al ciclo celular | Evita replicar el ADN fuera del momento correcto |
La cromatina merece una mención aparte: es ADN empaquetado alrededor de histonas. Esa organización protege el genoma, pero obliga a desmontar y reconstruir parte del empaquetamiento durante la replicación. En otras palabras, la célula no solo copia una secuencia; también mantiene la arquitectura que la hace legible.
Y cuando esa arquitectura o la corrección de errores se deterioran, el problema deja de ser teórico y entra de lleno en el terreno clínico.
Por qué este proceso importa en cáncer, diagnóstico y medicina personalizada
En biología médica, el interés no está solo en cómo se copia el ADN, sino en lo que pasa cuando la copia deja señales de inestabilidad. Los defectos en genes de reparación, la pérdida de proofreading o la presión replicativa crónica favorecen la acumulación de errores, y eso puede alimentar el desarrollo tumoral.
El estrés replicativo aparece cuando la horquilla de replicación se frena o se atasca por daños en el ADN, escasez de nucleótidos o interferencias estructurales. Si esto se repite, aumenta la probabilidad de roturas, reordenamientos y mutaciones. En cáncer, esa inestabilidad genómica no es un detalle secundario: muchas veces forma parte del problema de fondo.
También hay implicaciones directas para el diagnóstico. Algunas alteraciones en genes de reparación del ADN ayudan a explicar síndromes hereditarios de predisposición al cáncer, y la presencia de defectos en la reparación de desajustes puede dejar una firma molecular muy útil para decidir tratamientos. En medicina personalizada, ese tipo de información no se interpreta aislada: se cruza con la histología, los antecedentes familiares y el perfil genético completo del paciente.
Yo aquí suelo ser bastante pragmático: entender la replicación no sirve solo para aprobar biología. Sirve para leer mejor un informe oncológico, interpretar por qué una mutación importa más que otra y entender por qué dos tumores aparentemente parecidos no siempre responden igual.
Con eso en mente, la última pieza útil no es una definición más, sino un mapa mental que te permita quedarte con lo importante sin perderte en detalles.
La idea que más ayuda cuando pasas del esquema al informe genético
Si te quedas con una sola imagen mental, quédate con esta: una horquilla de replicación donde una enzima abre, otra coloca un cebador, la polimerasa alarga 5'→3' y varios sistemas vigilan el resultado. Yo uso ese mapa para leer mejor desde un manual de genética básica hasta un informe de secuenciación tumoral.
- Primero entiende la polaridad, porque explica por qué existe hebra líder y hebra rezagada.
- Después piensa en la corrección, porque ahí se decide si un fallo queda como lesión pasajera o como mutación estable.
- Por último añade el contexto celular: no es lo mismo una bacteria con cromosoma circular que una célula humana con cromatina y telómeros.
Si dominas esas tres capas, la replicación deja de ser una lista de enzimas y se convierte en una herramienta real para interpretar genética, enfermedad y diagnóstico molecular con bastante más criterio.