La duplicación del ADN bacteriano es uno de esos procesos que parecen simples hasta que se mira de cerca: en realidad, coordina decenas de proteínas, un control muy fino del tiempo de inicio y una logística molecular bastante elegante. En este artículo explico cómo se organiza la replicación en procariotas, qué hace cada componente de la maquinaria y por qué este mecanismo es tan importante en genética, microbiología y biomedicina.
Lo esencial de la copia del ADN bacteriano
- En bacterias, la replicación suele arrancar en un único origen, oriC, y avanza en dos direcciones.
- La apertura de la doble hélice depende de iniciadores como DnaA, helicasas, topoisomerasas y proteínas SSB.
- La síntesis nueva la realiza sobre todo la ADN polimerasa III, con una cadena líder y otra retardada.
- Los cebadores de ARN son imprescindibles para empezar, y luego se eliminan y sustituyen por ADN.
- La terminación no es solo “llegar al final”: también hay que resolver el entrelazamiento de las dos moléculas hijas.
- No todos los procariotas siguen exactamente el mismo patrón; en arqueas hay variantes que conviene no mezclar con el modelo clásico bacteriano.
Qué problema resuelve esta maquinaria celular
Cuando una célula procariota se divide, no puede permitirse copiar su genoma de manera improvisada. Necesita duplicar la información con rapidez, pero también con una fidelidad suficiente para que la descendencia conserve los genes que la hacen viable. Ahí está la clave: la replicación no es solo “hacer más ADN”, sino garantizar continuidad genética sin romper la lógica del genoma.
En bacterias, el modelo más estudiado es el de Escherichia coli: un cromosoma circular, un solo origen de replicación y dos horquillas que avanzan en sentidos opuestos. Esa simplicidad aparente ayuda a entender el proceso, pero no debe engañar al lector: incluso en este escenario, la célula tiene que abrir una doble hélice estable, mantener separadas las hebras, sintetizar nuevas cadenas, corregir errores y cerrar todo sin dejar nudos ni roturas. Yo suelo resumirlo así: copiar ADN no es el problema; copiarlo bien y a tiempo es lo difícil.Esta base te servirá para entender el resto del mecanismo, porque todo empieza en un punto concreto y termina con dos cromosomas funcionales. El paso siguiente es ver cómo se elige ese punto de arranque.
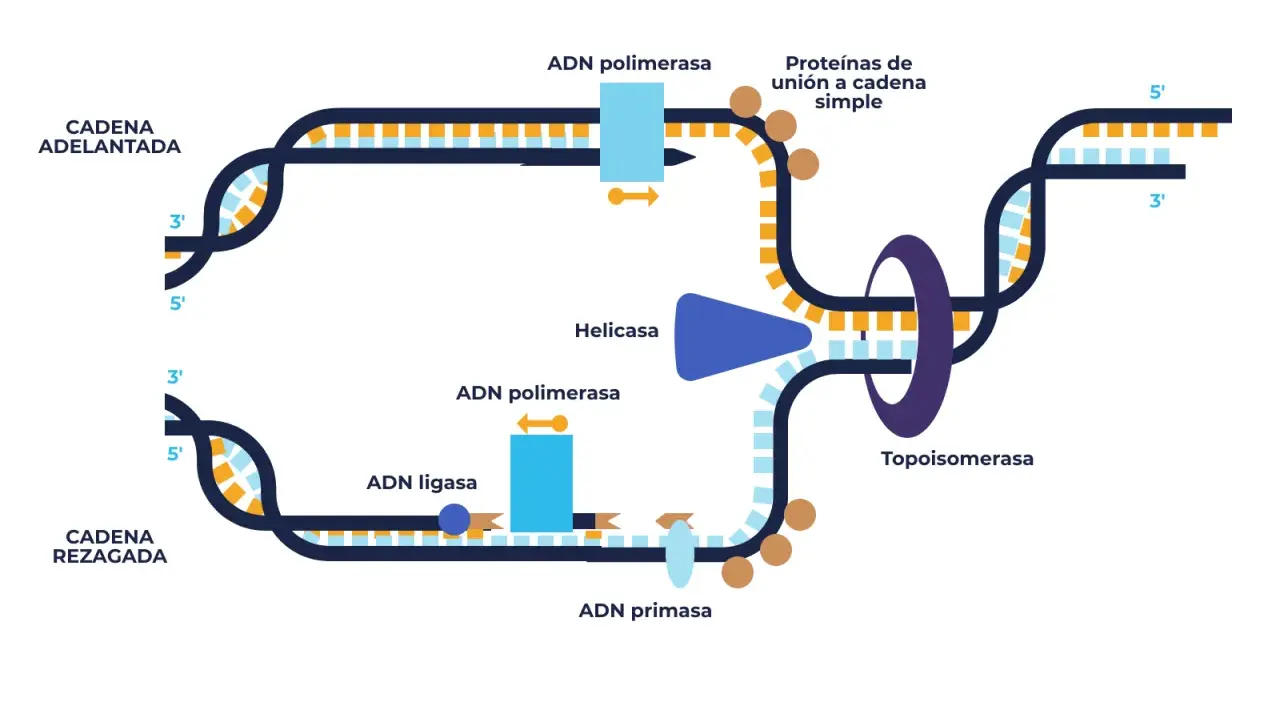
Cómo se inicia en el origen oriC
La replicación bacteriana arranca en una secuencia específica del ADN llamada oriC. En E. coli, este origen contiene regiones ricas en A-T, que se separan con más facilidad que las zonas ricas en G-C, y varios sitios de unión para la proteína iniciadora DnaA. Esa combinación no es casual: la secuencia funciona como una plataforma molecular que “autoriza” el inicio cuando la célula está preparada.
Primero se ensamblan moléculas de DnaA sobre oriC. Después, la región abierta por estas proteínas expone bases complementarias que facilitan el desenrollamiento inicial. A partir de ahí entran en juego otras piezas: la helicasa, que separa las hebras; el cargador de helicasa, que ayuda a colocarla correctamente; y las proteínas que estabilizan el ADN monocatenario para que no vuelva a cerrarse de inmediato. Si faltara este orden, la horquilla ni siquiera arrancaría con estabilidad.
Este punto de inicio también es el principal lugar de control del proceso. En bacterias, una vez montada la maquinaria, la célula ya no tiene tanto margen para “arreglar” la marcha; por eso la iniciación está tan regulada. Esa regulación explica por qué los errores en esta fase pueden alterar la velocidad de crecimiento o incluso favorecer inestabilidad genética. Y de ahí pasamos a la parte más útil para estudiar el tema: quién hace qué.
Qué proteínas hacen el trabajo pesado
En la replicación procariota no actúa una sola enzima, sino un conjunto muy coordinado. Para leerlo con claridad, conviene separar las funciones principales:
| Proteína o complejo | Función principal | Por qué importa |
|---|---|---|
| DnaA | Reconoce oriC e inicia la apertura local del ADN | Define dónde empieza todo |
| DnaB helicasa | Desenrolla la doble hélice | Permite que las hebras queden accesibles |
| DnaG primasa | Sintetiza cebadores cortos de ARN | Da un extremo 3' desde el que puede empezar la polimerasa |
| SSB | Estabiliza el ADN monocatenario | Evita que las hebras se reagrupe o formen estructuras secundarias |
| ADN polimerasa III | Extiende la nueva cadena de ADN | Es la principal replicasa bacteriana |
| β-clamp | Mejora la procesividad de la polimerasa | Permite copiar largas distancias sin que la enzima se desprenda |
| ADN polimerasa I | Elimina cebadores de ARN y rellena los huecos | Convierte los fragmentos iniciales en ADN continuo |
| ADN ligasa | Sella los enlaces finales entre fragmentos | Deja la cadena intacta y continua |
| Topoisomerasas | Alivian el superenrollamiento y resuelven el entrelazamiento | Evitan que la replicación se bloquee por tensión mecánica |
El detalle que más suele perderse en clase es este: la polimerasa no trabaja sola ni puede empezar desde cero. Necesita un cebador y un entorno físico controlado. Por eso, cuando se entiende el papel de las topoisomerasas, de la helicasa y de la primasa, todo encaja mejor. La maquinaria no solo copia; también mantiene el ADN en un estado operativo mientras lo hace.
Con esto ya se puede seguir la horquilla paso a paso, que es donde la replicación se vuelve realmente visible para quien estudia biología molecular.
Qué ocurre en la horquilla de replicación
La horquilla de replicación es la estructura en forma de Y donde se produce la síntesis nueva. Ahí las dos hebras originales se separan y se copian simultáneamente, pero no del mismo modo. La razón es química y estructural: las ADN polimerasas solo sintetizan en dirección 5' a 3', mientras que las dos hebras molde son antiparalelas. Esa asimetría obliga a organizar una cadena líder y una cadena retardada.
La cadena líder se sintetiza de forma continua, porque su orientación permite a la polimerasa avanzar de manera fluida conforme la helicasa abre el ADN. La cadena retardada, en cambio, se fabrica a saltos mediante fragmentos de Okazaki. Cada fragmento necesita un nuevo cebador de ARN, y después ese cebador se elimina, se reemplaza por ADN y se sella el corte. En bacterias, estos fragmentos suelen ser más largos que en eucariotas, del orden de aproximadamente 1000 a 2000 nucleótidos.Si lo miras con lupa, la horquilla funciona como una pequeña fábrica modular. La helicasa abre la vía, la primasa coloca el punto de arranque, la polimerasa III extiende la cadena y la β-clamp evita que la enzima se caiga al avanzar. Esa combinación de apertura, arranque y extensión explica por qué el proceso es tan rápido: en bacterias como E. coli, la horquilla puede avanzar a cientos de nucleótidos por segundo. Y no basta con avanzar; también hay que cerrar bien el proceso cuando las dos copias se encuentran.
Cómo se termina y cómo se separan las dos copias
La terminación no es simplemente el momento en que “ya no queda ADN por copiar”. En realidad, hay dos problemas distintos. El primero es detener la síntesis cuando las horquillas convergen. El segundo, igual de importante, es resolver el entrelazamiento físico de los dos cromosomas recién formados, que suelen quedar concatenados como anillos unidos.
En muchas bacterias, el final se alcanza cuando las dos horquillas se encuentran en una región opuesta al origen. En E. coli, además, existe el sistema Tus-Ter, que actúa como una especie de barrera direccional para ordenar la terminación. No todos los procariotas usan este mismo esquema, y conviene recordarlo para no generalizar en exceso a partir del modelo clásico. Después de la convergencia, entran en juego topoisomerasas como la topoisomerasa IV, que ayudan a separar físicamente las moléculas hijas.
Esto es más importante de lo que parece. Si la copia termina pero las moléculas quedan pegadas, la división celular falla o se vuelve errática. En otras palabras, la terminación no es un epílogo decorativo, sino una fase funcional que determina si el material genético podrá repartirse correctamente. A partir de aquí vale la pena comparar el modelo bacteriano con otros procariotas y con eucariotas, porque ahí aparecen las excepciones que más confunden.
Qué cambia en arqueas y por qué no conviene mezclarlo todo
Cuando se habla de procariotas, a menudo se piensa solo en bacterias, pero el grupo también incluye arqueas. Y aquí hay una advertencia útil para cualquiera que estudie genética molecular: no todos los procariotas copian su ADN exactamente igual. El esquema de una sola horquilla, un origen y una maquinaria bacteriana concreta describe muy bien a muchas bacterias, pero no a todos los procariotas por igual.
En arqueas, la replicación suele parecerse más a la eucariota en varios componentes: algunas usan múltiples orígenes, y sus proteínas de iniciación, helicasa y primasa comparten rasgos con las de células eucariotas. Eso no significa que sean “eucariotas pequeñas”, sino que la evolución ha conservado soluciones mixtas. Para mí, esta es una de las partes más interesantes del tema, porque rompe la idea simplista de que procariota equivale siempre a “modelo bacteriano estándar”.
La comparación también ayuda a estudiar mejor el caso bacteriano. Cuando se entiende qué es universal y qué es específico, se lee con más criterio cualquier esquema de libro o artículo. Y esa comparación se vuelve todavía más útil si el objetivo no es solo académico, sino también biomédico.
Por qué este proceso importa en genética y medicina
La replicación bacteriana no interesa solo por anatomía molecular. En genética, explica cómo se transmiten mutaciones, cómo se conservan plásmidos y cómo se mantiene la identidad de una población microbiana. En medicina, importa porque muchos antibióticos interfieren precisamente en esta maquinaria. Las fluoroquinolonas, por ejemplo, afectan a topoisomerasas bacterianas y alteran la copia del ADN, lo que frena la proliferación de la bacteria.
También tiene un peso enorme en resistencia antimicrobiana. Cuando una bacteria acumula cambios en genes relacionados con la replicación, la reparación o el mantenimiento del genoma, puede modificar su tasa de mutación o la eficacia con la que responde a ciertos fármacos. No siempre eso se traduce en resistencia directa, pero sí puede cambiar el escenario evolutivo en el que aparecen mutaciones útiles para la supervivencia. Ese matiz es importante: la replicación no “causa” por sí sola la resistencia, pero sí condiciona el terreno donde ocurre.
Desde una perspectiva más amplia, yo veo este proceso como una pieza puente entre biología básica y aplicación clínica. Entenderlo bien ayuda a leer mejor la microbiología, a interpretar mecanismos de acción de antibióticos y a comprender por qué algunas bacterias crecen tan rápido y se adaptan con tanta eficacia. Y, si lo estás estudiando, hay unas pocas ideas que conviene fijar antes de cerrar el tema.
Lo que conviene retener si quieres estudiar este tema con criterio
Si tuviera que dejar una versión muy limpia del proceso, diría esto: primero se selecciona un origen; después se abre la doble hélice; luego se monta la horquilla; más tarde se sintetizan una cadena continua y otra fragmentada; y, por último, se corrigen, se unen y se separan las moléculas hijas. Todo el proceso está pensado para que la célula gane velocidad sin perder precisión.
- Inicio no significa “empezar a copiar” sin más, sino reclutar una maquinaria específica en un origen concreto.
- El desenrollamiento del ADN genera tensión, así que las topoisomerasas no son accesorias: son necesarias.
- La polimerasa III es la protagonista de la elongación, pero depende de primasa, clamp, helicasa y SSB.
- La cadena retardada no es un error del sistema; es la solución obligada a la antiparalelidad del ADN.
- La terminación incluye la separación física de los cromosomas, no solo la última base copiada.
Si quieres memorizar la replicación en procariotas sin perderte en nombres, quédate con esa secuencia lógica y con dos ideas de fondo: la síntesis es semiconservativa y la maquinaria trabaja como un conjunto, no como enzimas aisladas. Ese es el punto en el que el proceso deja de ser una lista de proteínas y se convierte en un mecanismo comprensible.