Una guía rápida para leer los ejemplos mendelianos sin perderte en los cruces
- La herencia mendeliana describe rasgos controlados, en general, por un solo gen con dos alelos.
- La primera ley explica la uniformidad de la F1: si cruzas una línea pura dominante con una recesiva, toda la descendencia suele parecerse al progenitor dominante.
- La segunda ley muestra cómo reaparece el rasgo recesivo en la F2 con una proporción fenotípica típica de 3:1.
- La tercera ley solo funciona bien cuando los genes se segregan de forma independiente, algo que no siempre ocurre.
- En humanos, algunas enfermedades monogénicas encajan bien con este modelo, pero muchos rasgos cotidianos, como la estatura o el color de ojos, no lo hacen.
- Dominante no significa más frecuente ni más fuerte; significa que se expresa en heterocigosis.
Qué explican realmente las leyes de Mendel
Yo suelo empezar por una idea sencilla: las leyes de Mendel no describen “cómo se hereda todo”, sino cómo se comportan ciertos rasgos de herencia simple. En este modelo, cada carácter depende de un gen con dos alelos, uno aportado por cada progenitor, y el resultado visible se llama fenotipo, mientras que la combinación genética se llama genotipo. Cuando el alelo dominante enmascara al recesivo, el rasgo puede parecer muy fácil de predecir; ahí es donde Mendel acertó con tanta precisión.
Para orientarte rápido, esta tabla resume lo esencial de cada ley y el tipo de ejemplo que suele usarse para explicarla.
| Ley | Qué observa | Ejemplo clásico | Resultado esperado |
|---|---|---|---|
| Uniformidad | La F1 es homogénea cuando se cruzan líneas puras | AA x aa | 100% Aa, con fenotipo dominante |
| Segregación | Los alelos se separan al formar gametos | Aa x Aa | Genotipo 1:2:1 y fenotipo 3:1 |
| Distribución independiente | Dos genes distintos se heredan por separado | AaBb x AaBb | Fenotipos en proporción 9:3:3:1, si no hay ligamiento |
Ese esquema sirve como mapa mental, pero la comprensión real llega cuando lo llevas a un cruce concreto. Por eso el siguiente paso es ver el ejemplo clásico que hace visible la primera ley sin trucos ni saltos lógicos.
El ejemplo de los guisantes que hace visible la primera ley
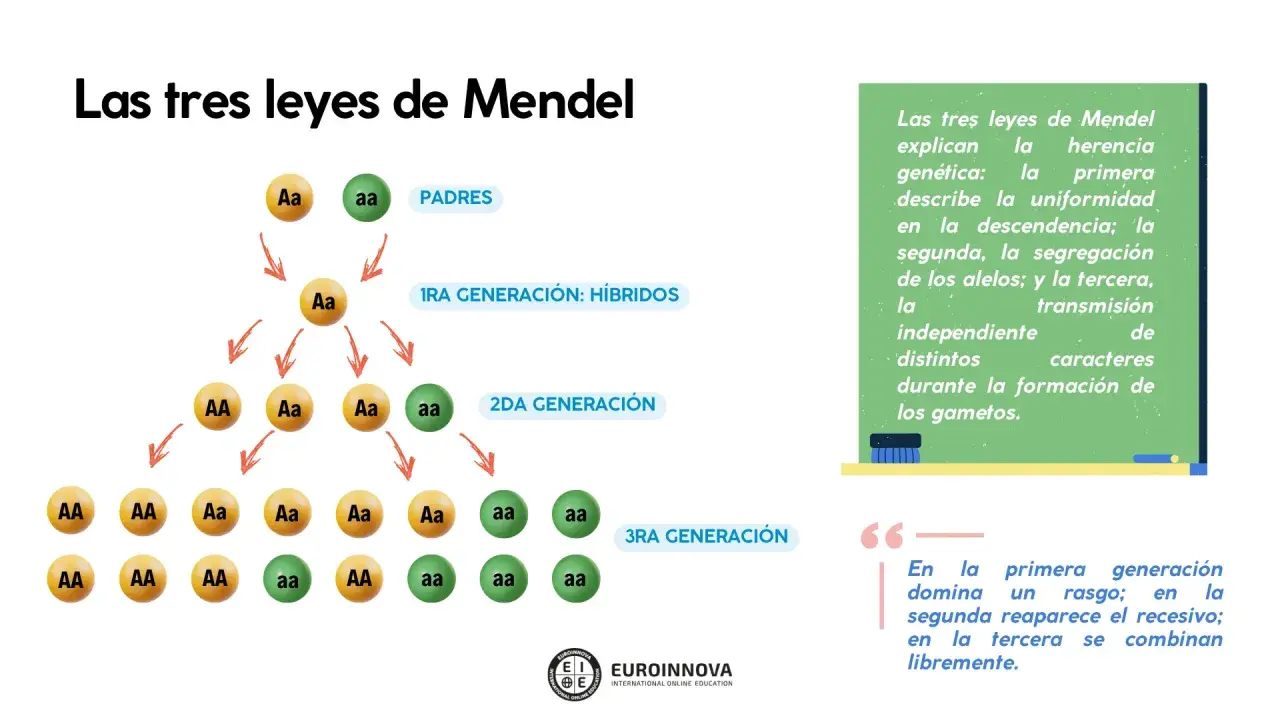
El ejemplo más didáctico es el de dos líneas puras de guisantes con rasgos opuestos, por ejemplo semillas amarillas y semillas verdes. Si cruzas una planta homocigota dominante, representada como AA, con una homocigota recesiva, aa, toda la descendencia de la primera generación filial será Aa. Fenotípicamente, todos mostrarán el rasgo dominante, porque ese alelo se expresa y el recesivo queda oculto.
Lo importante aquí no es memorizar letras, sino entender la lógica: en la F1 no aparece una mezcla intermedia, sino una descendencia uniforme. Ese detalle rompe una confusión muy extendida, porque mucha gente imagina que la herencia funciona como una dilución. Mendel demostró justamente lo contrario: los factores hereditarios se conservan y se combinan, no se “disuelven”.
Si yo tuviera que explicarlo en una sola frase, diría esto: un cruce entre líneas puras produce una F1 igual entre sí y parecida al progenitor dominante. A partir de ahí, ya se entiende por qué la segunda generación es tan reveladora y por qué el rasgo recesivo no desaparece, solo queda en silencio por un tiempo.
Cómo reaparece el rasgo recesivo en la segunda ley
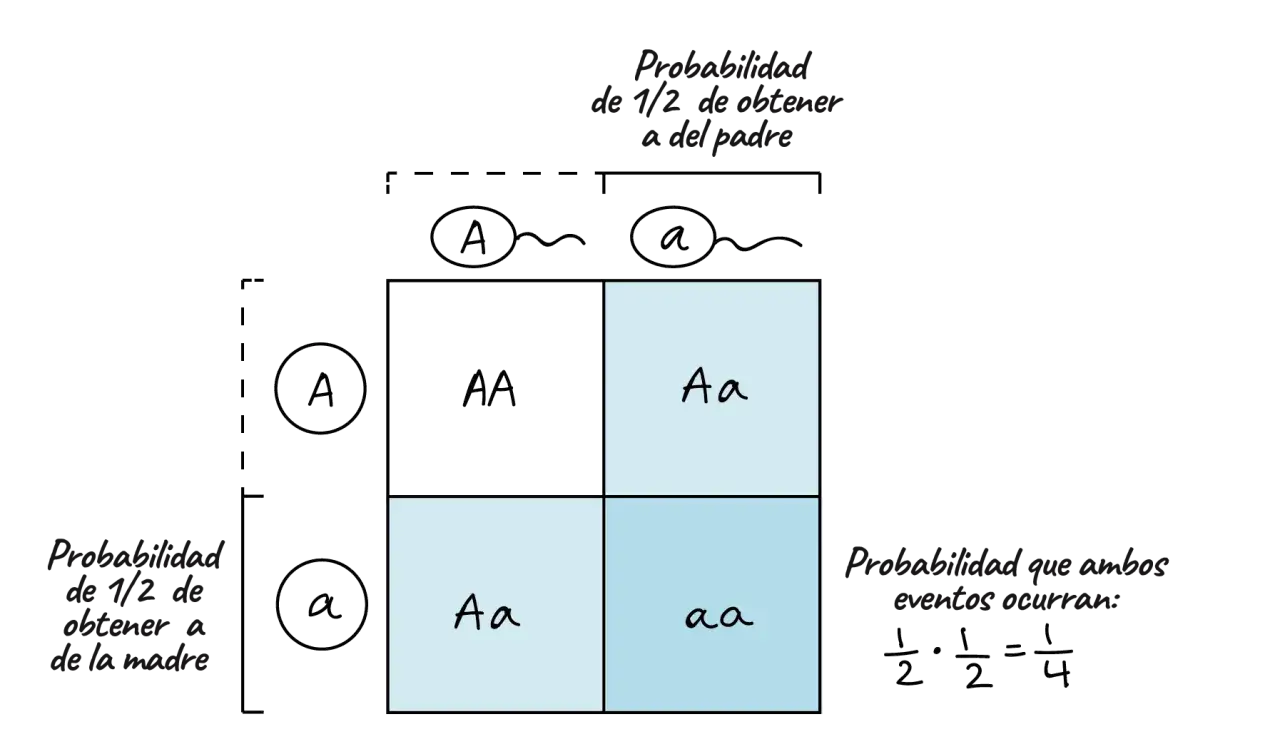
La segunda ley aparece cuando cruzas dos individuos de la F1, es decir, Aa x Aa. Aquí la cosa se vuelve más interesante, porque cada progenitor puede aportar dos tipos de gametos, uno con A y otro con a. El resultado genotípico es 1 AA : 2 Aa : 1 aa, y el resultado fenotípico suele ser 3 dominantes por 1 recesivo.
Yo insisto en este punto porque es donde más se cometen errores: la proporción no significa que “de cuatro hijos, uno saldrá sí o sí recesivo”. Significa que, en una gran cantidad de descendientes, la tendencia esperada se aproxima a esa relación. Cada embarazo es un evento independiente; haber tenido un hijo con un rasgo no cambia la probabilidad del siguiente.
En clase o en consulta, esta ley también ayuda a entender el concepto de portador. Un individuo heterocigoto puede no mostrar el rasgo recesivo, pero sí transmitirlo. Eso explica por qué una característica puede reaparecer después de saltarse una generación y por qué el árbol familiar, leído sin cuidado, puede engañar. Desde aquí ya puedes pasar a la tercera ley, donde el reto es seguir dos caracteres a la vez.
La tercera ley y los cruces dihíbridos
La tercera ley de Mendel, o ley de la distribución independiente, se ve cuando estudias dos rasgos al mismo tiempo, por ejemplo color y textura de la semilla. Si ambos genes se segregan de forma independiente, un cruce AaBb x AaBb puede producir cuatro tipos de gametos por progenitor y una descendencia fenotípica típica de 9:3:3:1. En el modelo clásico de los guisantes, eso permite obtener combinaciones como amarillas y lisas, amarillas y rugosas, verdes y lisas, o verdes y rugosas.La clave práctica está en la palabra independiente. Esta ley funciona bien cuando los genes están en cromosomas distintos o lo bastante separados en el mismo cromosoma como para recombinarse con frecuencia. Si están ligados, es decir, muy cerca entre sí, el patrón se altera y el 9:3:3:1 deja de ser una guía fiable. Ese matiz importa más de lo que parece, porque muchos errores de interpretación nacen de aplicar la tercera ley donde no toca.
En otras palabras, Mendel no “falló”; lo que pasa es que su modelo describe muy bien una parte de la realidad biológica, pero no toda. Y esa frontera es justo la que conviene conocer antes de llevar el esquema a ejemplos humanos.
Ejemplos humanos que sí encajan bastante bien
Si me pides ejemplos útiles en herencia humana, yo no empezaría por rasgos vistosos como el color de ojos, porque suelen ser demasiado complejos. Preferiría enfermedades o caracteres monogénicos, donde el patrón es mucho más limpio y la comparación con Mendel tiene verdadero valor. Ahí sí aparecen proporciones previsibles, sobre todo cuando hablamos de herencia autosómica dominante, autosómica recesiva o ligada al cromosoma X.| Patrón | Ejemplo frecuente | Qué suele verse en la familia | Probabilidad simple |
|---|---|---|---|
| Autosómica dominante | Acondroplasia, enfermedad de Huntington | Suele haber afectados en varias generaciones consecutivas | Si un progenitor es heterocigoto y el otro no está afectado, cada hijo tiene 50% de riesgo |
| Autosómica recesiva | Fibrosis quística, anemia falciforme | Puede saltarse generaciones; ambos padres suelen ser portadores | Si ambos padres son portadores, 25% afectado, 50% portador y 25% no afectado ni portador |
| Ligada al cromosoma X recesiva | Hemofilia A, daltonismo rojo-verde | Los varones suelen estar más afectados porque solo tienen un cromosoma X | Depende de si la madre es portadora o afectada |
Estos ejemplos son útiles porque conectan la teoría con la genética médica real. Ahora bien, también conviene decir algo que muchos materiales divulgativos omiten: rasgos como la estatura, la pigmentación general de la piel o gran parte de la variación del color de ojos no siguen un patrón mendeliano simple. Eso no los hace menos genéticos; solo significa que necesitan otro tipo de explicación.
Cuándo un ejemplo deja de ser mendeliano
Hay varios casos en los que el esquema clásico deja de ser suficiente, y yo los separo siempre porque cada uno rompe la predicción por una razón distinta:
- Herencia poligénica: intervienen varios genes a la vez, como ocurre con la estatura.
- Dominancia incompleta: el heterocigoto muestra un fenotipo intermedio, como en algunas flores rosadas.
- Codominancia: ambos alelos se expresan al mismo tiempo, como en el grupo sanguíneo AB.
- Ligamiento génico: dos genes cercanos en el mismo cromosoma tienden a heredarse juntos.
- Penetrancia y expresividad variables: el gen puede estar presente, pero no manifestarse igual en todas las personas.
También hay un error conceptual que merece una aclaración directa: dominante no significa más frecuente. Un alelo dominante puede ser raro, y uno recesivo puede ser común. Lo dominante se expresa en heterocigosis; eso es todo. Si separas esa idea de la frecuencia poblacional, dejas de arrastrar una confusión que aparece una y otra vez en genética básica.
Con estas limitaciones claras, ya se puede pasar a una lectura más madura de los árboles familiares y a una regla práctica que yo considero imprescindible antes de llamar “mendeliano” a un rasgo.
Qué conviene verificar antes de dar por mendeliano un rasgo
Cuando veo un ejemplo o una historia familiar, yo me hago cuatro preguntas antes de aceptar el patrón como mendeliano:
- ¿Parece depender de un solo gen, o hay señales de que intervienen varios?
- ¿El rasgo dominante realmente se comporta como dominante completo, o hay un fenotipo intermedio o doble expresión?
- ¿Los genes implicados parecen heredarse de forma independiente, o podrían estar ligados?
- ¿El patrón encaja mejor con herencia autosómica, ligada al X o con un modelo no mendeliano?
Si la respuesta a varias de esas preguntas es “no lo sé” o “no encaja del todo”, lo prudente es no forzar el modelo. En genética, simplificar demasiado suele ser peor que quedarse corto, porque crea expectativas erróneas sobre probabilidades, portadores y riesgo familiar. Mi regla práctica es esta: usa Mendel para entender el mecanismo básico, pero no para cerrar el caso cuando la evidencia apunta a un rasgo complejo.
Con esa frontera bien dibujada, los ejemplos dejan de ser ejercicios de memoria y pasan a ser una herramienta útil para leer herencia genética con más precisión. Ese cambio de perspectiva es, en la práctica, lo que más ayuda a interpretar una familia, una enfermedad o un rasgo sin caer en simplificaciones que luego no se sostienen.