Las leyes de Mendel siguen siendo la base más clara para entender cómo se heredan muchos rasgos biológicos, desde un carácter visible en plantas hasta ciertas variantes genéticas en humanos. En este artículo explico qué describen de verdad, cómo se leen sus cruces clásicos y en qué casos el modelo deja de ser suficiente. También verás por qué siguen importando hoy en genética clínica y en la interpretación de antecedentes familiares.
Lo esencial para entender la herencia mendeliana sin perderse
- Los rasgos se transmiten mediante genes con versiones alternativas llamadas alelos.
- La primera regla explica por qué la F1 puede salir uniforme cuando se cruzan líneas puras.
- La segunda regla describe cómo se separan los alelos al formar gametos y por qué aparece la proporción 3:1 en cruces simples.
- La tercera regla sirve para entender la herencia independiente de dos caracteres, con la proporción clásica 9:3:3:1.
- No todo encaja en el modelo clásico: dominancia incompleta, codominancia, ligamiento y rasgos poligénicos cambian el resultado.
- En clínica y medicina personalizada, estas reglas ayudan a leer patrones familiares, pero no sustituyen la interpretación experta.
Qué explican realmente estas reglas de la herencia
Yo suelo empezar por una idea sencilla: Mendel no describió toda la genética, sino un modelo predictivo muy potente para rasgos que se comportan de forma discreta. Ese modelo permite entender por qué un individuo hereda una combinación concreta de alelos y cómo esa combinación puede traducirse, o no, en un fenotipo visible.
Para no mezclar conceptos, conviene separar cuatro niveles básicos. El gen es la unidad hereditaria; el alelo es cada una de sus versiones; el genotipo es la combinación de alelos que tiene un individuo; y el fenotipo es el resultado observable de esa combinación, influido también por el contexto biológico.
| Concepto | Qué significa | Por qué importa |
|---|---|---|
| Gen | Fragmento de información hereditaria asociado a un rasgo o función. | Es la unidad básica que se transmite de padres a hijos. |
| Alelo | Una versión concreta de un mismo gen. | Explica por qué un mismo rasgo puede presentarse de maneras distintas. |
| Genotipo | La combinación de alelos que porta un individuo. | Ayuda a predecir probabilidades de herencia. |
| Fenotipo | La manifestación observable del rasgo. | Es lo que vemos, medimos o diagnosticamos. |
| Heterocigoto y homocigoto | Dos alelos distintos o dos alelos iguales para un gen. | Determinan cómo se expresa un rasgo en un cruce. |
La clave, en la práctica, es no confundir dominancia con “mejor” o “más frecuente”. Dominante solo significa que un alelo puede imponerse en la expresión del fenotipo en ciertos contextos; no dice nada sobre su valor biológico. Con esta base clara, ya se entienden mejor las tres reglas una por una.
La primera ley y la uniformidad de la F1
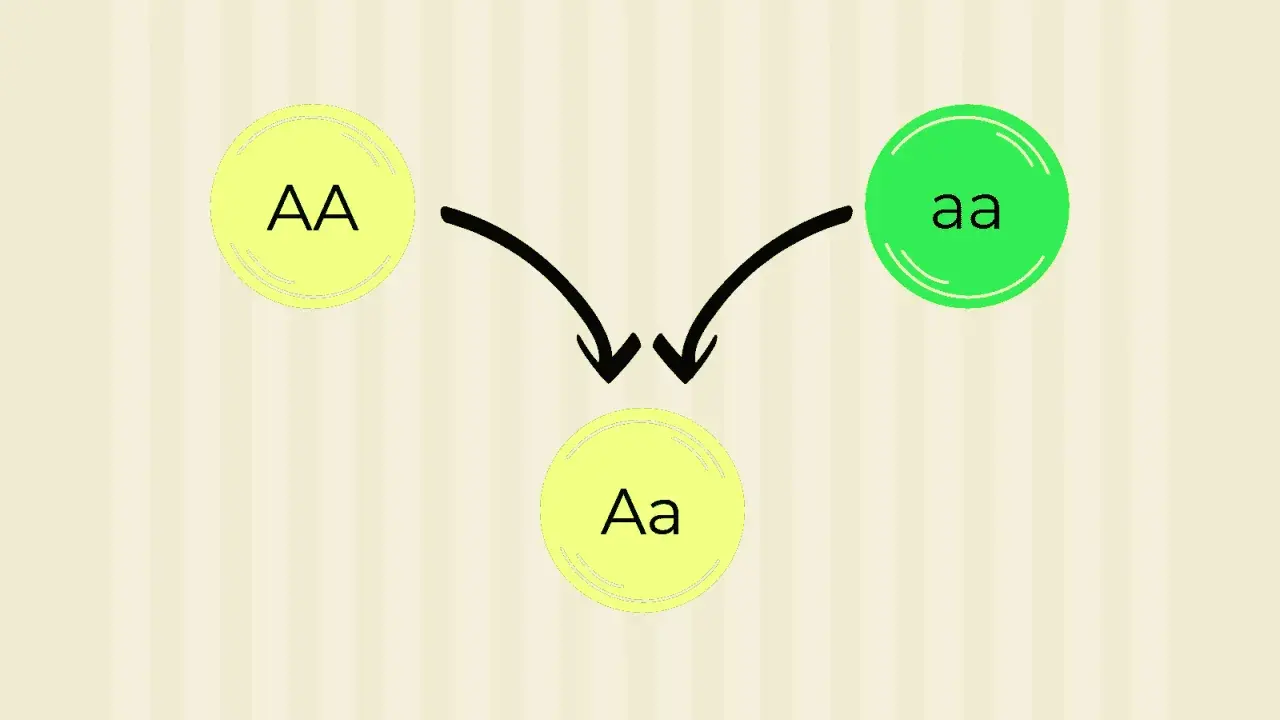
La primera ley describe lo que ocurre cuando cruzo dos líneas puras para un carácter concreto. Si una planta es homocigota dominante y la otra homocigota recesiva, toda la primera generación filial, la F1, resulta uniforme. En el caso más clásico, un cruce AA x aa produce descendencia Aa en todos los individuos.
Eso se traduce en una idea muy útil: si el alelo dominante tiene dominancia completa, toda la F1 muestra el fenotipo dominante, aunque genéticamente sea heterocigota. En los guisantes de Mendel, este principio permitió ver que la apariencia externa no siempre revela la composición genética real.
- Los progenitores aportan un alelo cada uno.
- La F1 suele ser genéticamente uniforme.
- El fenotipo dominante enmascara al recesivo si hay dominancia completa.
La segunda ley y la segregación de los alelos
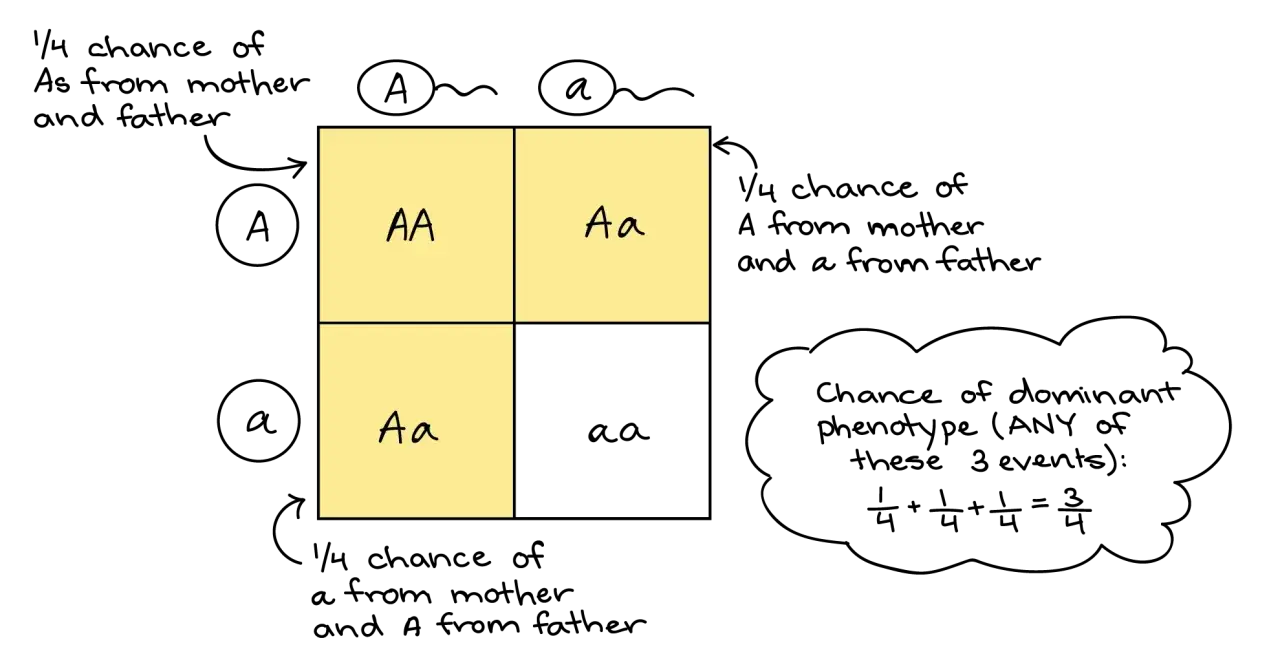
La segunda ley explica que, cuando se forman los gametos, los dos alelos de un par se separan. Dicho de otro modo: cada gameto recibe solo uno de los dos alelos. Esa idea permite predecir qué combinaciones aparecerán en la descendencia cuando dos heterocigotos se cruzan.
Si cruzo Aa x Aa, la distribución genotípica esperada es 1 AA : 2 Aa : 1 aa. Si además hay dominancia completa, el reparto fenotípico pasa a ser 3 dominantes : 1 recesivo. Esta es la proporción clásica que casi todo el mundo memoriza, pero lo importante no es la cifra, sino el mecanismo que la produce.
Yo la leo así: la segregación ocurre antes de la fecundación, no después. El alelo no “decide” en el embrión; ya viene separado desde el gameto. Ese detalle explica por qué un rasgo recesivo puede reaparecer en la F2 aunque haya desaparecido en la F1.
| Cruce | Resultado genotípico | Resultado fenotípico | Qué enseña |
|---|---|---|---|
| Aa x Aa | 1 AA, 2 Aa, 1 aa | 3 dominante, 1 recesivo | Los alelos se separan al formar gametos. |
Esta ley es especialmente útil cuando quiero estimar probabilidades de transmisión en rasgos monogénicos. En cuanto el carácter depende de varias variantes o aparece un tipo de dominancia distinta, la proporción deja de ser tan limpia.
La tercera ley y la distribución independiente
La tercera ley añade una capa más: cuando observo dos caracteres distintos, los alelos de un gen se distribuyen de forma independiente respecto a los del otro, siempre que no estén ligados y que el contexto biológico lo permita. Es el principio que explica por qué heredar el color de una semilla no obliga a heredar al mismo tiempo su forma.
En un cruce dihíbrido clásico, como AaBb x AaBb, la F2 suele mostrar la proporción 9:3:3:1. Ese patrón surge porque cada pareja de alelos se reparte por separado en los gametos. La mezcla de combinaciones posibles aumenta de forma notable, y ahí se ve la potencia real del modelo de Mendel.
Lo interesante es que esta ley no habla solo de “más variedad”, sino de independencia estadística entre caracteres. Si un rasgo condiciona otro, o si ambos genes están muy próximos en el mismo cromosoma, la proporción esperada deja de cumplirse con precisión.
Por eso esta ley es muy útil como punto de partida, pero no debe interpretarse como una regla universal para cualquier combinación de rasgos. A partir de aquí ya toca mirar las excepciones, que en genética real son mucho más frecuentes de lo que suele parecer en clase.
Dónde dejan de bastar en genética real
Si uno quiere usar este modelo con rigor, tiene que saber dónde falla. No porque Mendel se equivocara, sino porque estudió un tipo de herencia muy concreto, especialmente limpio, con rasgos discretos y relaciones alélicas simples. La biología real es bastante más variada.
| Patrón | Qué cambia respecto al modelo clásico | Ejemplo orientativo |
|---|---|---|
| Dominancia incompleta | El heterocigoto muestra un fenotipo intermedio. | Flores rosas entre rojas y blancas. |
| Codominancia | Ambos alelos se expresan a la vez. | Grupo sanguíneo AB. |
| Alelos múltiples | En la población hay más de dos versiones del mismo gen. | Sistema ABO. |
| Ligamiento genético | Genes cercanos en el mismo cromosoma no se heredan de forma independiente. | Rasgos situados en regiones próximas del cromosoma. |
| Herencia poligénica | Muchos genes contribuyen al mismo rasgo. | Estatura, pigmentación o riesgo cardiometabólico. |
| Penetrancia y expresividad variables | El mismo genotipo no produce siempre el mismo grado de manifestación. | Algunas variantes asociadas a enfermedad familiar. |
Aquí aparece una precisión que me parece esencial: tener una variante no equivale siempre a expresar el rasgo de forma predecible. La penetrancia indica cuántos portadores muestran el fenotipo; la expresividad señala cuánto se manifiesta. Es una diferencia pequeña en apariencia, pero enorme en asesoramiento genético.
En rasgos complejos, además, el ambiente pesa. Nutrición, edad, sexo biológico, epigenética y otros factores modulan la expresión final. Por eso la genética moderna no sustituye el modelo mendeliano: lo amplía. Y esa ampliación es justo lo que importa cuando se trabaja con medicina personalizada.Qué aporta hoy a la genética clínica y a la medicina personalizada
En clínica, este modelo sigue siendo muy útil para leer árboles familiares, estimar riesgos de transmisión y decidir qué tipo de estudio genético tiene sentido pedir primero. Cuando una enfermedad sigue un patrón autosómico dominante, autosómico recesivo o ligado al sexo, el análisis mendeliano sigue siendo una herramienta de primera línea.
Pero yo no lo usaría nunca como una sentencia. Una predicción genética buena no se basa solo en un gen, sino en el contexto del caso: historia familiar, edad de inicio, penetrancia conocida, pruebas disponibles y calidad de la información clínica. Esa mirada evita errores frecuentes, como confundir predisposición con diagnóstico o simplificar un rasgo complejo como si fuera binario.
También hay una dimensión ética que no conviene ignorar. Cuando una variante genética se relaciona con enfermedad, la conversación con el paciente o la familia debe separar con claridad probabilidad, certeza y limitación técnica. Eso vale tanto para una prueba diagnóstica como para una estimación de riesgo reproductivo.
En la práctica, el valor de este modelo no está en prometer respuestas absolutas, sino en ofrecer una estructura para pensar mejor la herencia. Y esa estructura sigue siendo muy potente, precisamente porque permite distinguir entre lo que se puede predecir con bastante solidez y lo que exige más matices clínicos.
Lo que conviene recordar antes de interpretar una herencia familiar
Si tuviera que dejar una idea final, sería esta: Mendel no explica toda la genética, pero sí enseña a pensar con orden. Primero identifico si el rasgo parece monogénico; después compruebo si hay dominancia completa, ligamiento o participación de varios genes; y solo entonces intento calcular probabilidades con cierta confianza.
Cuando ese filtro se aplica bien, las reglas mendelianas dejan de ser una lista para memorizar y pasan a ser una forma rigurosa de leer la herencia. Y, en la práctica, esa es la diferencia entre una explicación superficial y una interpretación realmente útil.
Por eso, más que repetir de memoria las leyes de Mendel, merece la pena entender cuándo funcionan, cuándo solo orientan y cuándo hay que pasar a un análisis genético más amplio y más cuidadoso.