Las ligasas de ADN son las enzimas que cierran las “costuras” químicas del genoma: unen fragmentos separados y restauran el esqueleto fosfodiéster de la molécula. Sin ellas, la replicación dejaría huecos, la reparación quedaría incompleta y muchas rutas de mantenimiento del ADN perderían eficiencia. En este artículo explico qué hacen, cómo funcionan, en qué procesos participan y por qué su estudio importa tanto en biología molecular como en medicina.
Lo que conviene tener claro sobre las ligasas de ADN
- Su función principal es sellar roturas monocatenarias y unir extremos compatibles de ADN.
- No añaden información nueva: la secuencia ya tiene que estar alineada antes de que actúen.
- Necesitan un 3'-OH y un 5'-fosfato; sin esa química, no hay ligación eficiente.
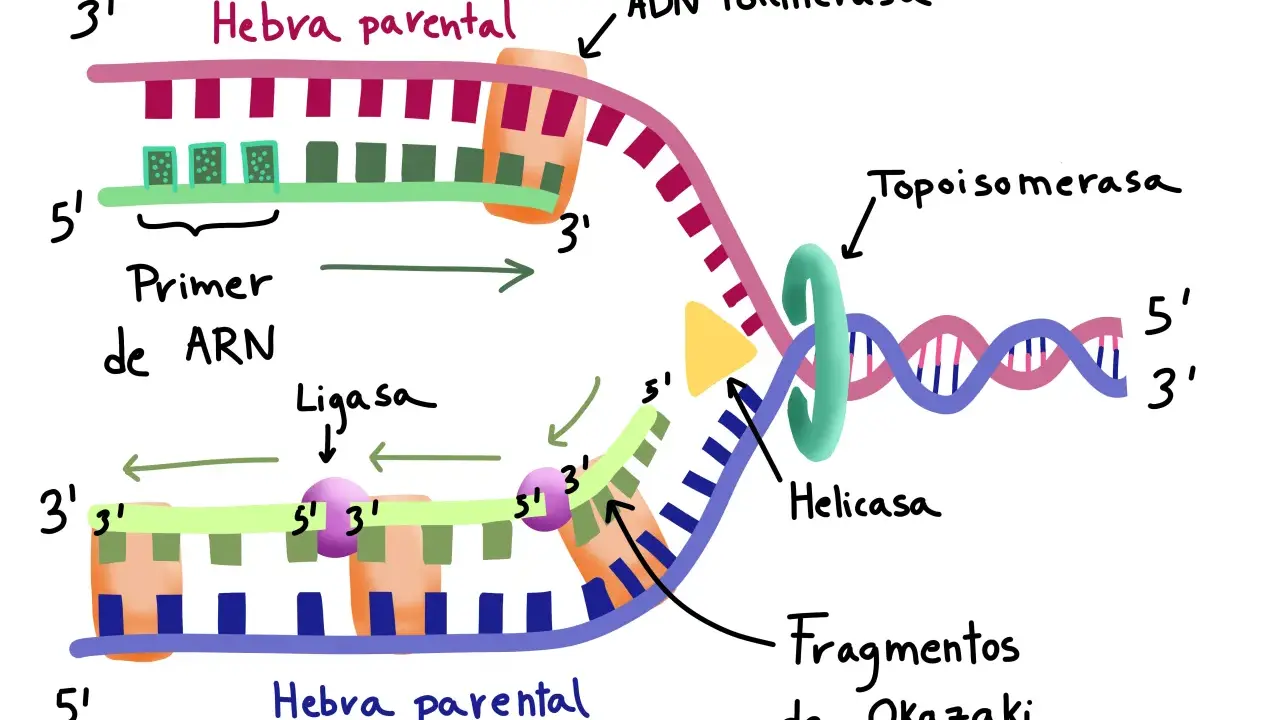
- Trabajan en replicación, reparación y recombinación, especialmente en la maduración de fragmentos de Okazaki.
- En eucariotas predominan las ligasas dependientes de ATP; en muchas bacterias, las dependientes de NAD+.
- Si fallan, aumenta la inestabilidad genómica y pueden aparecer fenotipos clínicos relevantes.
Qué hace una ligasa de ADN y por qué no es una polimerasa
Yo la describo como la enzima que termina el trabajo que otras ya iniciaron. La polimerasa construye la cadena; la ligasa la sella. Esa diferencia parece pequeña, pero es decisiva: una polimerasa añade nucleótidos siguiendo una plantilla, mientras que una ligasa solo puede cerrar un hueco químico ya preparado.
Su diana es una ruptura muy concreta, el nick: un corte en una sola hebra donde quedan enfrentados un extremo 3'-OH y un extremo 5'-fosfato. Si esos grupos no están presentes y correctamente alineados, la enzima no tiene nada que “pegar”. Por eso me gusta insistir en que la ligasa no es un adhesivo universal, sino una herramienta de precisión.
Con esa idea clara, el siguiente paso es entender la química exacta que usa para hacer la unión.
Cómo cierra un nick paso a paso
La reacción de ligación consume energía y, en términos simples, convierte dos extremos compatibles en un enlace fosfodiéster continuo. En las ligasas de ADN más estudiadas, esa energía procede de ATP; en muchas bacterias, de NAD+. Además, Mg2+ suele ser un cofactor esencial para que la catálisis avance con eficiencia.
- Adenilación de la enzima. La ligasa forma un intermediario ligasa-AMP en su sitio activo.
- Transferencia del AMP al ADN. El AMP pasa al extremo 5'-fosfato del nick y “activa” ese extremo.
- Formación del enlace final. El 3'-OH vecino ataca al fosfato activado y se forma el enlace fosfodiéster.
- Liberación del AMP. La molécula queda sellada y la ligasa puede reiniciar otro ciclo.
En extremos cohesivos la reacción suele ser más eficiente, porque las bases complementarias ayudan a alinear las hebras; en extremos romos, la geometría es peor y la ligación se vuelve más exigente. Esa limitación explica muchos resultados irregulares en el laboratorio y también ayuda a entender por qué la célula organiza tan bien sus rutas de reparación.
Esa organización se ve con mucha claridad cuando miramos en qué procesos celulares se apoya de verdad esta enzima.
En qué procesos celulares es imprescindible
En la célula, la ligasa no actúa de forma aislada. Normalmente entra al final de una cadena de eventos en la que otras enzimas han copiado, recortado o rellenado el ADN. Donde más peso tiene es en tres contextos: replicación, reparación y recombinación.
Replicación
En la hebra retardada, la síntesis es discontinua y genera fragmentos de Okazaki. En eucariotas suelen medir alrededor de 100-200 nucleótidos; en bacterias, con frecuencia, 1.000-2.000 nucleótidos. Una vez retirados los cebadores de ARN y rellenados los huecos, la ligasa sella la cadena para que el genoma quede continuo.
Reparación
También es clave en rutas que corrigen daños frecuentes, como la reparación por escisión de bases y por escisión de nucleótidos. Después de que nucleasas y polimerasas hagan su parte, queda un último nick que debe cerrarse. Si ese paso falla, la célula acumula roturas de una sola hebra que pueden convertirse en roturas de doble hebra durante la replicación.
Lee también: ADN y genes - La guía para entender la molécula de la vida
Recombinación y roturas de doble hebra
En la vía de unión de extremos no homólogos, la ligasa IV trabaja junto con proteínas accesorias para volver a conectar extremos rotos. Aquí la exigencia es alta: la célula necesita reparar rápido, pero sin introducir demasiada inestabilidad. Esa tensión entre velocidad y fidelidad es una de las razones por las que la reparación del ADN es tan relevante en genética y en oncología.
Si miramos qué enzima hace cada variante de este trabajo, aparecen diferencias importantes entre bacterias y eucariotas.
Qué tipos de ligasas conviene distinguir
No todas las ligasas de ADN se comportan igual. La distinción más útil, en mi opinión, es combinar el cofactor que usan con el contexto biológico en el que trabajan. Esa lectura evita confusiones cuando se comparan bacterias, levaduras y células humanas.
En humanos, los genes más citados son LIG1, LIG3 y LIG4. No codifican una única solución, sino especializaciones distintas para problemas distintos.
| Tipo | Cofactor | Contexto habitual | Función destacada | Qué recordar |
|---|---|---|---|---|
| Ligasas dependientes de ATP | ATP | Eucariotas y arqueas | Replicación y reparación | Son las más relevantes en células humanas. |
| Ligasas dependientes de NAD+ | NAD+ | Muchas bacterias | Sellado de nicks durante replicación y reparación | Sirven como contraste útil en biología comparada. |
| Ligasa I | ATP | Núcleo | Maduración de fragmentos de Okazaki y reparación | Es la gran ligasa de la replicación. |
| Ligasa III | ATP | Núcleo y mitocondria | Reparación de roturas monocatenarias y mantenimiento mitocondrial | Importa mucho en mantenimiento genómico. |
| Ligasa IV | ATP | Núcleo | Unión de extremos no homólogos | Es la más ligada a roturas de doble hebra. |
Este mapa es útil porque ayuda a leer artículos, protocolos y resultados genéticos sin mezclar funciones que no son equivalentes. Y precisamente en el laboratorio se nota mucho cuándo una ligasa hace bien su trabajo y cuándo el sistema completo está mal preparado.
Cómo se aprovecha en el laboratorio y qué suele salir mal
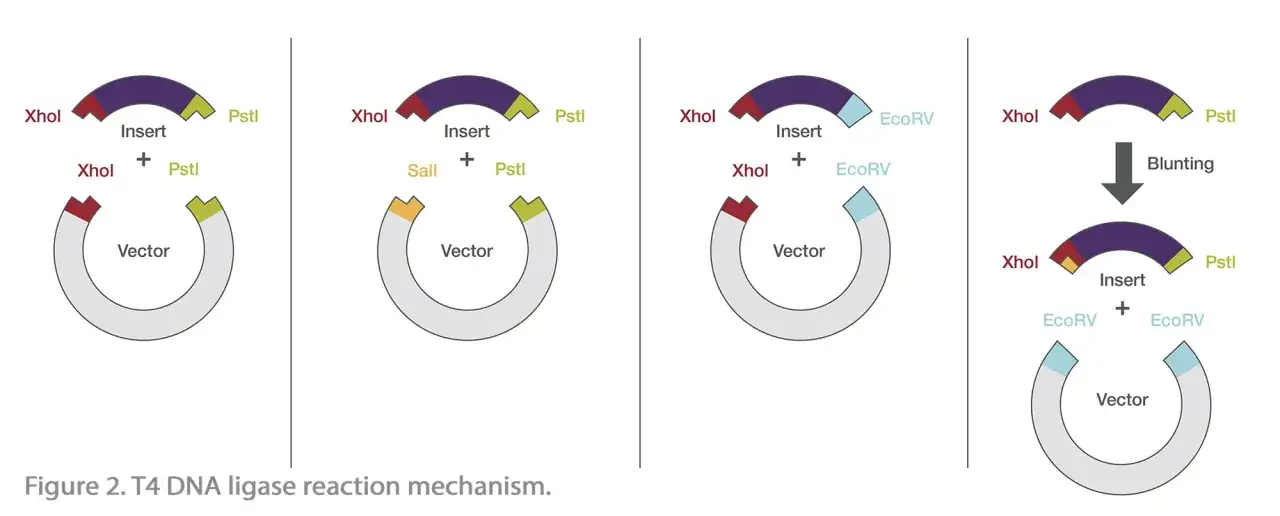
En biología molecular, la ligasa es una de esas enzimas que parecen sencillas hasta que algo falla. Se usa para clonar insertos, unir adaptadores en bibliotecas de secuenciación, reparar extremos después de un corte enzimático y cerrar construcciones de ADN diseñadas a medida. Cuando el resultado no sale, casi nunca es porque “la ligasa no funciona”; suele ser un problema de diseño, de extremos incompatibles o de preparación deficiente del ADN.
- Extremos incompatibles: si el insert y el vector no tienen geometría o química compatibles, la ligación cae mucho.
- Falta de fosfato en 5': sin ese grupo no se forma el enlace final.
- ADN sucio: sales, fenol, etanol o guanidinio pueden frenar la reacción.
- Vector auto-ligado: si no se controla, el vector puede cerrarse sobre sí mismo y reducir el rendimiento útil.
- Expectativas irreales: los extremos romos ligan, pero lo hacen peor y exigen más paciencia.
Yo suelo resumirlo así: la ligasa no compensa un mal diseño experimental, solo remata un diseño bien hecho. Esa idea también sirve para entender por qué las alteraciones en ligasas humanas tienen consecuencias biológicas tan serias.
Qué pasa cuando falla y por qué importa en medicina
Cuando una ligasa no funciona bien, el problema no es solo un enlace mal cerrado. Se acumulan roturas, aparecen errores de replicación y la célula entra en un terreno de inestabilidad genómica. En términos clínicos, eso puede traducirse en síndromes raros de reparación del ADN, inmunodeficiencia, alteraciones del desarrollo o mayor sensibilidad a la radiación, especialmente cuando el defecto afecta a la ligasa IV o a rutas muy dependientes de ligación.
En particular, LIG4 es relevante porque la misma maquinaria participa en la recombinación V(D)J, así que las variantes patogénicas pueden afectar la maduración del sistema inmune. En genética clínica conviene ser prudente: una variante en un gen de ligasa no equivale automáticamente a enfermedad. Hay que mirar el contexto, la zigosidad, el tipo de variante, la evidencia funcional y el fenotipo del paciente. Esa es una de las razones por las que la interpretación de variantes en medicina personalizada exige más que leer un informe genético en diagonal.
También hay una lectura oncológica importante. Si una célula tumoral tiene alteradas rutas de reparación del ADN, su respuesta a radioterapia o a fármacos que dañan el ADN puede cambiar. No siempre eso se traduce en una decisión clínica directa, pero sí en una pista biológica que merece atención.
Con ese panorama, la pregunta útil ya no es solo qué hace la enzima, sino qué conviene recordar al interpretar resultados o protocolos.
Lo que conviene recordar al interpretar datos sobre reparación del ADN
La idea más importante es simple: la ligasa no corrige la secuencia, solo sella el ADN preparado por otras enzimas. Si faltan el extremo 3'-OH, el 5'-fosfato o la alineación correcta, la reacción se detiene antes de empezar. Por eso el contexto químico pesa tanto como la enzima en sí.
- Si lees un protocolo, comprueba primero el estado de los extremos y no solo la marca comercial de la ligasa.
- Si revisas un estudio genético, separa bien la función de LIG1, LIG3 y LIG4; no son intercambiables.
- Si trabajas con clonación o bibliotecas, recuerda que la limpieza del ADN y la compatibilidad de extremos suelen explicar más fallos que la propia reacción de ligación.
- Si analizas variantes clínicas, interpreta siempre la ligasa dentro de la ruta de reparación completa, no como una pieza aislada.
En mi lectura, las ligasas de ADN son una buena prueba de cómo la biología molecular depende de pasos pequeños pero decisivos: copiar, procesar y, al final, cerrar sin dejar cicatrices innecesarias.